
Este artículo es un breve extracto del tomo III de la Enciclopedia del cáncer, dedicado el metabolismo tumoral.
Es un artículo muy útil para saber qué “puntos de presión” podremos pulsar con acciones prácticas, que luego aplicaremos entre todos en la red de Oncología Metabólica.
En él, comprenderemos que los matices deben presidir siempre nuestro análisis y que las hipótesis no son nunca amenazas. Al contrario, veremos que la hipótesis del efecto Warburg reverso puede explicar algunas aparentes paradojas y completar (antes que sustituir) al efecto Warburg “convencional”.
***
Este es un tema espinoso, que enerva a muchos miembros del panorama “alternativo”, pero que debe abordarse si queremos ser exhaustivos, porque varios procesos fisiológicos y neoplásicos encajan (en cierta manera) en ese modelo.
El efecto Warburg reverso abre la puerta a un “modelo parasitario del cáncer”, donde las células tumorales ejercen una especie de “pastoreo” del entorno que las rodea. Como veremos, no es un modelo ajeno a las relaciones metabólicas en otras células sanas, y ya se vislumbró esa posibilidad cuando analicé la autofagia tumoral.
El problema es que también abre la posibilidad de que el tumor sea capaz de usar cetonas y ácidos grasos con fines energéticos, algo que solivianta a muchos científicos que promulgan que el cáncer es una enfermedad metabólica.
En primer lugar, no entiendo por qué las hipótesis pueden soliviantar a nadie. Ser científico implica ser implacable con las ideas: no enamorarse de ellas sino usarlas de forma instrumental, como mero medio para hallar la verdad, asesinándolas sin compasión si surgen otras mejores. Las hipótesis de trabajo deben circular con total libertad, luchando entre ellas acalorada e intensamente. Son el medio para encontrar recursos prácticos y clínicos de calidad.
En segundo lugar: como recuerdo en el blog y en los libros, una cosa es la posibilidad de utilizar cetonas y ácidos grasos con fines catabólicos, energéticos, y otra que el tumor sigue ligado al consumo de combustibles como glucosa y glutamina para la obtención de material anabólico que sustente su elevada proliferación, y ese siempre ha sido su talón de Aquiles.
Cuando se dice que hay que “matar de hambre” al tumor, no se usa una correcta analogía: es muy difícil matarlo de hambre, porque es sencillo para él conseguir energía de múltiples formas, gracias a su plasticidad y recursos. Su auténtico punto débil son los materiales anabólicos que, cuando se le niegan y se añaden otros ataques en otros puntos de presión metabólicos, constituye un hecho a veces catastrófico.
Analicemos por tanto la evidencia sin prejuicios, con mente abierta y con el único fin de obtener soluciones prácticas que ayuden a los enfermos, no que aposenten nuestras creencias. En caso contrario estaríamos haciendo lo mismo que todos los patanes que se aferran a mantras manidos y que tratan de desestimar todo lo que los refuta.
Introducción al efecto Warburg reverso
Antes de empezar, un vídeo donde hago un resumen del contenido de este artículo:
Un estudio publicado el año 2009 por un grupo de investigadores de la Universidad de Filadelfia, al frente del cual se encontraba Michael P. Lisanti, supuso un enorme revuelo.
En el estudio se proponía que las células epiteliales cancerígenas inducían un efecto Warburg en las células asociadas del estroma, en concreto fibroblastos deficientes en una proteína llamada caveolin-1 (CAV-1, que analizo en el tomo III de la enciclopedia del cáncer) y que, una vez diferenciados en miofibroblastos, basarían su metabolismo en la fermentación de la glucosa en vez de en fosforilación oxidativa y producirían diferentes metabolitos “de desecho”, como lactato, que se eliminaría a través de un MCT-4 (transportador monocarboxilado-4) y podrían ser utilizadas como sustratos metabólicos por las células tumorales a través de un MCT-1.
Es decir, que el efecto Warburg estaría limitado a las células del estroma, mientras que las tumorales seguirían conservando intacta su capacidad oxidativa mitocondrial, algo que hasta ahora se consideraba imposible.
En el libro analizo en profundidad el estroma tumoral y sus componentes no celulares y celulares (entre los que se encuentran fibroblastos y miofribroblastos), porque son de importancia capital, sea cierto o no el efecto Warburg reverso.

Es decir, se produciría una especie de ‘pastoreo’, donde los fibroblastos serían literalmente ‘ordeñados’ por las células tumorales en una relación casi parasitaria. Las implicaciones de este hecho serían enormes, porque supondría una diferenciación metabólica radical entre ambas zonas del microambiente tumoral: la propiamente cancerígena y las células y proteínas no neoplásicas, de apoyo, componentes del estroma. Y el efecto Warburg ya no sería una característica propia de las células tumorales, sino de las adyacentes, supuestamente no patológicas.
Las consecuencias no se detendrían aquí: implicaría además que las mitocondrias de las células tumorales serían plenamente funcionales y capaces de oxidar todo tipo de combustibles metabólicos, entre los que se encontraría el lactato de desecho procedente de los fibroblastos. El mismo grupo publicó al año siguiente un estudio donde se afirmaba que las cetonas y el lactato fomentaban el crecimiento tumoral y las metástasis (estudio).
El nuevo concepto atentaba contra muchas asunciones que hasta entonces se habían solidificado en el panorama ‘alternativo’, entre ellas que las células tumorales no podían usar lactato como combustible, que en realidad era un resultado de su metabolismo fermentativo, un residuo de la actividad metabólica de las neoplasias.
También abría la puerta a considerar que la betaoxidación era posible en las células tumorales, aunque ya hemos visto, en el tomo III, que no es necesario que el efecto Warburg reverso sea cierto para que ya sea posible que las células tumorales puedan oxidar ácidos grasos de cadena muy larga en los peroxisomas (y quizá en algunos casos en mitocondrias hasta cierto punto funcionales) para obtener energía.
Las reacciones a los estudios no se hicieron esperar: tanto Thomas Seyfried como algunos blogueros especializados en la divulgación de las bondades de las dietas ‘low carb’ o cetogénica publicaron artículos denostando la metodología empleada para alcanzar las conclusiones del artículo, entre ellas que se habían inyectado directamente cetonas a los ratones del estudio, lo cual implicaba una situación anómala y no fisiológica.
Yo mismo descarté en mi blog las conclusiones del segundo estudio, asumiendo que las fallas metodológicas no podían contradecir tantos estudios acumulados que afirmaban lo contrario, pero con ello sólo estaba dejando que mis sesgos (y mi ignorancia) tomaran las riendas. Sigo sin estar convencido de que el efecto Warburg reverso haya demostrado ser un modelo cierto, pero desde luego hay argumentos y resultados que obligan a reflexionar y estoy obligado a sopesar sus pros y sus contras y también cómo podría encajar en el efecto Warburg “convencional” sin chirriar.
Pros de la hipótesis del efecto Warburg reverso
El principal: es un proceso YA existente en procesos fisiológicos no patológicos
El cáncer es un enemigo flexible y tenaz, que usará todas las herramientas a su alcance para sobrevivir. Y dichas herramientas son aproximadamente las mismas que las de un embrión o una herida e incluso similares a las emprendidas por una agrupación de microbios. Un cáncer no puede inventarse caminos metabólicos y bioquímicos que el cuerpo no tenga ya disponibles en su arsenal.
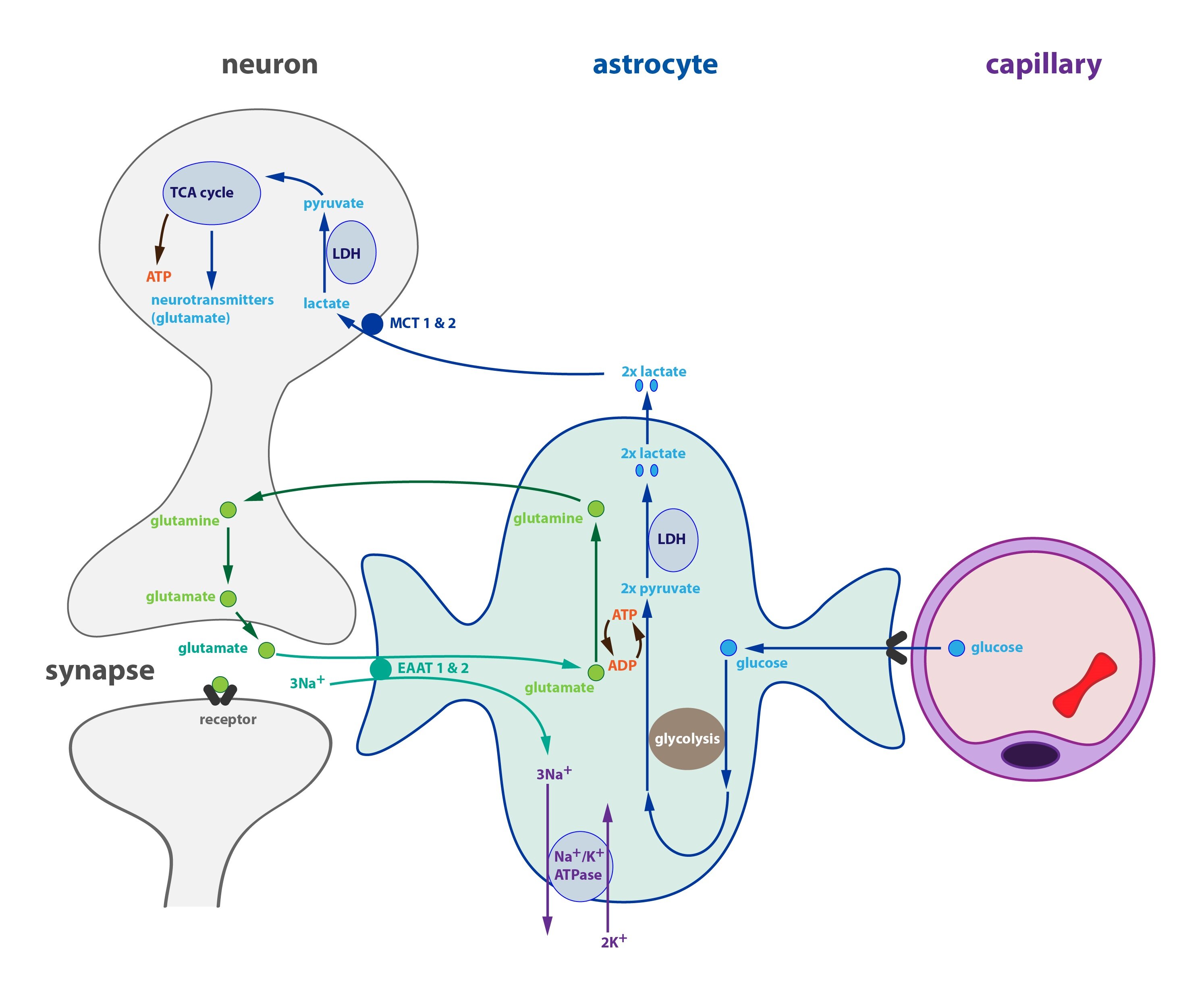
En el cerebro hay dos tipos principales de células, las neuronas y las células gliales. Las primeras realizan el ‘trabajo’ auténtico que identificamos como propio del cerebro. Las segundas son células de soporte metabólico que nutren a las primeras y atienden a sus necesidades. Es un sistema donde unas células “sirven” metabólicamente a otras. No es una relación parasitaria, porque astrocitos y oligodendrocitos no se sacrifican por las neuronas, pero sí ejercen una función de suministradoras de combustibles. La cuestión es: ¿Cuáles son dichos combustibles?
En casi cualquier tratado de neurología se repite como un mantra que “el combustible preferido por el cerebro es la glucosa”, y en algunos incluso podemos leer afirmaciones aún más tajantes: “el cerebro sólo puede usar glucosa”.
Ambas afirmaciones son ciertas si nos restringimos a un ámbito de alimentación como la occidental, donde el ayuno está prácticamente prohibido y los niveles de carbohidratos de la dieta hacen imposible que el cerebro use nada que no sea la omnipresente glucosa. Pero que nuestro estilo de vida promueva la glucosa no significa necesariamente que sea el preferido ni el principal combustible metabólico cerebral.
Nuestro cerebro ha sido moldeado por millones de años durante los cuales el escenario en latitudes con fuertes cambios estacionales (no así en zonas tropicales y ecuatoriales) era muy diferente: los cuerpos cetónicos eran habituales combustibles debido al ayuno y los alimentos grasos y proteicos, y la glucosa un ocasional regalo que aparecía estacionalmente cuando la primavera y el verano ponían a disposición alimentos con mayor carga de carbohidrato.
***
NOTA: no identifico a los carbohidratos con el demonio. Hay poblaciones humanas (por ejemplo, los habitantes de Kitava, isla del pacífico que fueron estudiados debido a su óptima salud metabólica), cuya latitud pone a su disposición abundantes carbohidratos y su alto consumo no conduce a problemas metabólicos, porque se inscriben en un contexto coherente, de igual forma que es coherente que un inuit consuma sobre todo grasas y proteínas y apenas carbohidratos. Kitavans e inuits están igualmente sanos porque se alimentan de forma coherente a su latitud. Analicé ese tema en este artículo.
***
El cerebro ha sido ‘educado’ por la evolución para ‘saltar al modo glucosa’ cuando puede, para almacenar energía en forma de panículo adiposo. Si hay glucosa disponible el cerebro la usará porque ‘cree’ que ese estado será poco duradero. Pero en el mundo actual no hay escasez de glucosa, algo que el cerebro no ‘sabe’: siempre hay glucosa disponible, por eso es siempre la que utiliza, creyéndose en un (perpetuo) estado de “abundancia que no durará”. La glucosa no es el combustible preferido, sino el más habitual en los tiempos modernos (artículo).
Con ese escenario, no es extraño que en publicaciones como ésta se siga afirmando que es la glucosa el combustible cerebral preferido.
No obstante, lo que llama la atención es que la glucosa nutre preferiblemente a astrocitos y oligodendrocitos (células gliales de soporte metabólico, que alimentan a las neuronas), y que usan la glucólisis exacerbada, fermentativa, para producir lactato que, a su vez, parece ser el alimento preferido por las neuronas. Si ‘la glucosa es el combustible preferido por las células gliales’, entonces, ‘el lactato es el preferido por las neuronas’. Pero este hecho puede provenir, repito, del modo como nos alimentamos.
Este hecho, observable en cualquier cerebro sano alimentado con dieta occidental, tiene gran relevancia porque, de manera similar, las células cancerígenas (según el modelo clásico basado en el efecto Warburg) muestran una tendencia a la fermentación y producen un exceso de lactato como producto de desecho. Además, el ciclo de la glutamina-glutamato se usa de forma inversa tanto en neoplasias como en astrocitos. Analizo todos esos factores metabólicos en el tomo III de la enciclopedia del cáncer, dedicado al metabolismo tumoral.
{kind=link}
Los tumores cerebrales suceden preferiblemente en las células gliales (astrocitomas y glioblastomas en astrocitos; oligodendrogliomas en oligodendrocitos). Los gliomas son devastadores cánceres con abrumadoras tasas de glucólisis, y parece lógico que comiencen allí si la glucólisis de los astrocitos preneoplásicos está ya exacerbada. Es decir, si siempre se alimentan de glucosa para nutrir el ciclo de la fermentación que produce lactato, ya están manteniendo un tipo de metabolismo muy parecido al de las neoplasias.
Por tanto, este comportamiento observado entre astrocitos y oligodendrocitos (que son los que habitualmente se convierten en neoplásicos en tumores cerebrales) y neuronas, típico de una dieta occidental, guarda similitudes con la hipótesis del modelo parasitario del cáncer, que describiremos a continuación.
Sin embargo, parece que los astrocitos y neuronas no sólo pueden ser alimentados también con cuerpos cetónicos hepáticos en vez de con glucosa, sino que los propios astrocitos pueden ser fuentes de síntesis de cetonas (estudio). No sólo eso, sino que su metabolismo ejercería importantes funciones neuroprotectoras, invalidando la teoría de que ‘el cerebro sólo puede usar glucosa’ e incluso la de que ‘la glucosa es el combustible preferido por el cerebro’. Nada más lejos de la realidad (estudio, estudio).
En el tomo III analizo también la enorme influencia del lactato en el cáncer y la teoría del “lactate shuttle”. Su papel corrobora hasta cierto punto el modelo parasitario del cáncer, cuya hipótesis describo en este artículo, pero debe quedar claro que otros estudios demuestran que las neuronas ‘no prefieren’ usar lactato, sino que no tienen más remedio que utilizarlo ante el exceso de glucosa que nutre a los astrocitos y que a su vez las nutre a ellas (estudio).
Según el modelo del “lactate shuttle”, el lactato no sólo se usaría como combustible por otros órganos, bien tras el paso intermedio de la gluconeogénesis hepática, o bien mediante la oxidación directa en la célula que lo produce, sino que podría usarse en intercambios célula-célula: el lactato que es metabolito de desecho de una célula, será el combustible oxidativo para otra célula próxima. Esa teoría sostiene el metabolismo glucolítico entre neuronas y células gliales, del que acabamos de hablar, pero también explicaría el llamado ‘efecto Warburg reverso’, que analizamos en este artículo (estudio, estudio).
La existencia del efecto Warburg reverso podría explicar la dificultad de tratar el cáncer de forma universalmente eficaz, incluso con terapias metabólicas. Si fuera cierto, estaríamos en un serio aprieto, porque las células tumorales pasarían a ser metabólicamente “normales”, similares a lo que sucede entre astrocitos y neuronas alimentadas con dietas occidentales llenas de carbohidratos.
Contras de la hipótesis del efecto Warburg reverso
1-La correlación entre proliferación y fermentación está muy estudiada
Y se produce en todo tipo de tejidos que necesitan crecer: sean células del sistema inmunitario que deben multiplicarse para hacer frente a una amenaza o lo que sucede tras un daño tisular o la proliferación típica de un embrión.
Las neoplasias crecen en función de su inutilización del aparato OXPHOS y de la asunción de un metabolismo fermentativo. La correlación entre producción de lactato y agresividad está muy estudiada. Asumir que sólo los fibroblastos y las células del estroma fermentan implicaría que sólo el tejido estromal crece y no así las células tumorales, algo que no se ha observado. Para proliferar, hay casi una obligatoriedad de usar fermentación en vez de respiración. Por tanto, la lucha contra la fermentación (sea en las células del estroma o en las tumorales) sigue siendo una necesidad, exista o no el efecto Warburg reverso.
Nota: es cierto que la masa tumoral está constituida en su mayor parte por estroma no tumoral, y que las células estromales del tumor crecen a un ritmo superior a las que forman parte de los tejidos sanos.
2-En etapas avanzadas, la hipoxia que se establece en amplias zonas del tumor hace que las células tumorales no puedan oxidar combustibles
Y no pueden hacerlo porque carecen del sustrato para ello, por lo que deberán usar igualmente la fermentación para avanzar, un proceso que produce lactato como desecho.
En ese caso, la maquinaria oxidativa no tendría sentido porque no pueden fermentarse ácidos grasos ni cuerpos cetónicos. Como las células tumorales más agresivas sobreviven en entornos hipóxicos, hiperproliferativos y angiogénicos, seguimos necesitando combatir la fermentación, aunque podamos reconocer la posibilidad de que las células neoplásicas sean más flexibles metabólicamente de lo que habíamos pensado al suponer que siempre se producía el efecto Warburg en ellas.
¿Podemos conciliar el efecto Warburg y el efecto Warburg reverso?
Quizá sí.
Propongamos una interesante hipótesis que conciliaría ambas posturas: en etapas iniciales el tumor podría mostrar características de efecto Warburg reverso, con crecimiento del tejido estromal y respiración normal de las células tumorales, que no crecerían tan rápido. Como ya hemos visto, un tumor benigno se relaciona con mayor fibrosis, producto de la actividad excesiva de las células estromales, y los tejidos fibróticos tienen mayor tendencia a desembocar en neoplasias malignas.
Vimos también que la fibrosis induce cambios tensionales que erosionan la homeostasis mecánica y que pueden condicionar metabólica y proliferativamente a las células, dificultando además la oxigenación, nutrición y eliminación de desechos metabólicos de las células inmersas en ese estroma.
En ese caso la proliferación de fibroblastos podría inducir una hipoxia en las células del parénquima que, a su vez, podría desembocar en una fermentación y una proliferación de las células epiteliales, y las consecuencias de ese metabolismo (acidez extracelular, angiogénesis exacerbada, etc) se traducen, a todos los efectos, en una neoplasia.
El efecto Warburg reverso implicaría, por tanto, tal vez, asumir que el inicio del cáncer podría suceder en el material estromal, en alguna alteración previa de los fibroblastos o de la producción aberrante de colágeno por parte de estas células y que la transformación neoplásica podría ser una consecuencia de este hecho, que desembocaría en un efecto Warburg “normal”.
Es decir, el efecto Warburg reverso podría inducir un efecto Warburg ‘ortodoxo’ en las células epiteliales, lo cual permitiría conciliar ambas ideas en vez de descartarlas como opuestas. Tal vez por esa razón, los inhibidores de glucólisis serían efectivos no sólo en células tumorales de zonas hipóxicas, sino en fibroblastos deficientes de CAV-1 (estudio).
Además de que la proliferación del estroma fermentativo puede conducir a una hipoxia y a un efecto Warburg en las células epiteliales, que se transforman definitivamente en tumorales, algunos estudios proponen, además, un mecanismo de realimentación desde las células tumorales que conservan su OXPHOS hacia las estromales fermentativas, basado en los ROS, generados profusamente desde las mitocondrias de las células epiteliales del parénquima tumoral.
Los ROS inducen un estrés oxidativo que facilita la expresión de factores como el HIF-1 (factor de crecimiento por hipoxia), que induce una sobreexpresión de autofagia y degradación lisosomal, lo cual conlleva mayor pérdida de CAV-1 y mayor tendencia a la proliferación y la agresividad estromal.
En el artículo dedicado a la autofagia tumoral estudiamos la aparente paradoja de que tanto la inhibición de las últimas etapas como la potenciación de la autofagia son acciones antitumorales. Tal vez se deba a que ambos hechos contrapuestos se producen en “escenarios” diferentes: uno en el estroma (donde la inhibición de la autofagia es antitumoral) y otro en las células tumorales propiamente dichas (donde es la exacerbación de la autofagia lo que produce efectos antitumorales).
A su vez, el resultado de esa autofagia y fermentación exacerbadas en el estroma implica la entrega a las células epiteliales de nutrientes. Tal vez ésa sea la razón por la cual funcionan tanto los inhibidores de últimas etapas de autofagia como los promotores de autofagia: los primeros inhibirían la autofagia estromal, impidiendo que produjera elementos nutritivos para las células tumorales, y los segundos favorecerían una autofagia en las células epiteliales del tumor, que podría correlacionar con eventos que las condujesen a la muerte.
El círculo vicioso de realimentación entre ambos escenarios del microambiente está servido, y estroma y parénquima “conversan” en un camino de aceleración mutua que llevaría al desastre (estudio).
Puede existir además una realimentación del lactato producido en zonas tumorales hipóxicas con alta fermentación hacia zonas normóxicas que conservan su capacidad oxidativa y que lo usarían como combustible metabólico (estudio).
Los estudios que hablan del efecto Warburg reverso, y que han comenzado a aflorar desde 2009, se ven desde entonces desde otra perspectiva: ya no suponen una ‘amenaza’ a la ortodoxia del efecto Warburg, sino otro punto de vista que en realidad lo certifica: puede que el inicio de la fermentación se produzca en los fibroblastos y esa ola de crecimiento afecte a las epiteliales en forma de hipoxia y que, al fermentar igualmente, también proliferen.
Esta hipótesis de inicio de la carcinogénesis se añade a otras también probables, como los estudios que parecen confirmar la existencia de defectos mitocondriales en las células tumorales y que sugieren que ésa puede ser también una razón de inicio de una carcinogénesis.
Algunos tumores NO surgen a partir de una cicatriz fibrótica previa, así que podemos suponer que el origen del cáncer puede ser variado y múltiple, aunque al final acabe por conducir al mismo punto: la hiperproliferación fermentativa.
Como quiera que sea, los estudios que parecen confirmar hasta cierto punto la existencia de fermentación estromal han abundado desde entonces (estudio, estudio, estudio, estudio). Algunos de ellos llegan a afirmar que no hay constancia de que las células tumorales usen la glucólisis fermentativa, que estaría reservada para los fibroblastos, confirmando la validez del efecto Warburg reverso (estudio). Si eso fuera cierto, la prueba PET (tomografía de emisión de positrones) mediría entonces el metabolismo fermentativo del estroma tumoral, no de las células tumorales.
Otros hablan de una situación de endosimbiosis y la utilización del lactato proveniente de los fibroblastos por parte de las células endoteliales convertiría la masa tumoral en una asociación de organismos unicelulares. La dispersión de las células en una malla estromal tumoral que se infiltra entre ellas, convierte esta hipótesis en plausible y puede explicar otras características que las asemejan a microorganismos como levaduras. El canibalismo de las células en este estado, pastoreando y engullendo a células de soporte estromal parece confirmarlo (estudio).
Volveremos a abordar este tema en el importante apartado dedicado a la biofísica del cáncer: los cambios metabólicos se producen a la vez (en una correlación donde resulta casi indiscernible identificar causas y consecuencias) que los cambios físicos bioquímicos y estructurales, que a su vez implican cambios en la polaridad de las membranas y modificación de los campos electromagnéticos (estudio).
Los microtúbulos y sus oscilaciones tendrán mucho que ver, y volveremos a estudiarlos en el apartado dedicado a la mente (si estos temas te suenan a ‘magufada’ te aconsejo que revises tus prejuicios y esperes a leer los argumentos que los explican científicamente y las hipótesis razonables que a partir de ellos construiremos).
Aunque el escepticismo ante la supuesta universalidad del efecto es muy alto, con razón, los datos convierten las acciones sobre la matriz extracelular tumoral y sus componentes celulares en objetivos terapéuticos muy atractivos. Otros estudios proponen que se use el supuesto efecto Warburg reverso para encontrar terapias basadas en él (estudio).
Esas técnicas irían dirigidas a eliminar la ayuda mutua que se establecería entre células tumorales y estroma (ROS en un sentido, que forzarían autofagia estromal; entrega de nutrientes en otro, que alimentarían a la célula tumoral). Hablaremos de algunas de ellas extensamente en el proyecto dedicado por excelencia a plantear acciones prácticas: Oncología Metabólica.
En definitiva: puede que, algunas veces, en etapas iniciales del cáncer, cuando ’sólo’ el estroma presenta efecto Warburg (reverso), los defectos mitocondriales se produzcan en las células del estroma tumoral como los fibroblastos, y que su actividad conduzca a una posterior glucólisis en las células del parénquima.
Y puede suceder, por tanto, que, en esas etapas iniciales de algunos cánceres, las células tumorales puedan consumir lactato e incluso cuerpos cetónicos, pero su procedencia será local, de la actividad metabólica del estroma, no del organismo. Una vez malignizado por completo, es decir, cuando las células tumorales proliferan debido a su recién adquirido fenotipo fermentativo, el efecto Warburg se establece totalmente. Por lo tanto, sigue teniendo vigencia y NO implica que la dieta cetogénica ni el resto de medidas metabólicas dejen de ser eficaces.
No veo motivos, según esa hipótesis, para que cambie nuestro planteamiento frente a las células tumorales más agresivas, que demuestran un fuerte efecto Warburg, pero debemos ampliar los enfoques terapéuticos hacia las mitocondrias de las células estromales, que adquieren una importancia tan grande o aún mayor que las propias células tumorales.
Por eso trato el tema del estroma tumoral con enorme profundidad en el tomo III de la enciclopedia del cáncer y será un tema estrella a tratar en Oncología Metabólica.
Libros imprescindibles para caminar hacia un paradigma realmente útil y eficaz
La «Enciclopedia del Cáncer» es una serie de 7 libros revolucionarios. El recurso definitivo para aquellos que buscan una comprensión profunda de esta enfermedad compleja. Te lleva a un viaje de conocimiento sin precedentes que cambiará la forma en que entiendes esta enfermedad.
Si buscas opciones de tratamiento más allá de los enfoques convencionales, la «Enciclopedia del Cáncer» te presenta una amplia gama de terapias innovadoras que mejoran el pronóstico y ofrecen esperanza real a los enfermos de cáncer. Estas páginas contienen valiosas herramientas y estrategias que pueden marcar la diferencia en la lucha contra esta enfermedad.
A partir de la hipótesis de que todos los cánceres son una única enfermedad metabólica, analiza los “puntos débiles” específicos que comparten y que los diferencia de los tejidos sanos y propone terapias prácticas combinadas que podrían mejorar considerablemente la cantidad y calidad de vida de los enfermos.
Explorando el cáncer desde múltiples puntos de vista, estos libros lo analizan con una perspectiva metabólica, desentrañando los misterios que rodean su origen y desarrollo. Descubre cómo el sistema inmunológico, la microbiota, el metabolismo, las emociones y la biofísica están interconectados en la batalla contra el cáncer.
La precompra incluye pertenencia a grupo privado que comparte experiencias y resultados. Y dará acceso durante un año al proyecto Oncología Metabólica, una revolución terapéutica colaborativa: una comunidad pionera en el diseño empírico colectivo de terapias combinadas con un enfoque metabólico, que mejorará radicalmente el tratamiento.
Al precomprar el pack de libros te ahorrarás casi un 70% respecto de la compra individual de cada libro (cada libro sale de esa forma a menos de 9 euros).
Super interesante. Yo tengo un camcer en el peritoneo 1 de 6 millones de mujeres lo tiene. Salí sorteada . Pero mi actitud para enfrentar y afrontar la enfermedad me está curando me daban 3 meses de vida . Llena de tumores malignos en todo mi abdomen
2 cirugías y 6 quimioterapias con homeópata que me mandó dieta cetogenica ciertas vitaminas , no harinas no carbohidratos y casi nada de azucar y menos proceaada y me siento explendida . Llevo 10 meses de tratamiento. Me dieron de vacaciones 15 días donde comí pastas helados pizzas y cuando me hicieron hace 15 días los análisis todo salió bien menos el tumoral que salió elevado. Así que empecé de nuevo a mi rutina a mis jugos verdes a comer mucha proteína semillas frutos secos ciertas frutas solo 3 y pocas cantidades , verduras cocidas mucho brocoli zapallito italiano cebolla pimientos. Mañana después de 15 días de diera me harán otro análisis y me harán un petscan para determinar si vuelco a las quimioterapias por vena o vía oral . Sin dejar la parte holistica combino los 2 , a pesar que sé que al ponerme tanto veneno a mi cuerpo por la quimioterapia tengo neuropatia en mis pies. El alma debe estar tranquilo y tener paz y dar amor siempre.. la vida es preciosa y no debemos desperdiciarla con pensamientos negativos ,debemos hacer actividad fisica y vivir en paz. Besos para todos
Gracias, Roxy, espero que todo te vaya bien
Muchísimo ánimo Roxy, toda la fuerza y energía positiva. Gracias por compartirlo
Alfonso, en cierta manera al hilo del efecto Warburg Iñigo San Millán acaba de publicar el siguiente estudio:
Role of Lactate in the Regulation of Transcriptional Activity of Breast Cancer-Related Genes and Epithelial-to-Mesenchymal Transition Proteins: A Compassion of MCF7 and MDA-MB-231 Cancer Cell Lines
https://www.biorxiv.org/content/10.1101/2023.03.23.533060v2
Por si es de tu interés
Un abrazo
Gracias, Faruk,
EL trabajo de Íñigo es interesantísimo
Otro abrazo.
Muy interesante articulo, gracias por compartir
Gracias, Jose Luis
Poca gente tiene claro que cáncer se ayuda de otro tipo de células que no son cancerosas, como criados.
Me acuerdo de un artículo de hace unos años del 2012 que decía:
En la nueva investigación, Nelson y sus colaboradores descubrieron que un tipo de célula no cancerosa, y normal -el fibroblasto- que se encuentra cerca del cáncer, daña el ADN y estimula el crecimiento del cáncer, cuando es expuesto a la quimioterapia. Bajo circunstancias normales, los fibroblastos ayudan a mantener la integridad estructural del tejido conectivo, y juegan un papel crítico en la curación de heridas y la producción de colágeno.
En concreto, la quimioterapia causa que los fibroblastos produzcan una proteína llamada WNT16B, cerca del tumor, y los altos niveles de esta proteína permiten que las células cancerosas crezcan e invadan el tejido circundante, resistiendo, así, a la quimioterapia.
https://www.infosalus.com/actualidad/noticia-mecanismo-hace-cancer-resistente-quimioterapia-podria-conducir-desarrollo-terapias-mas-eficaces-20120805190827.html
El cáncer como dices es sumamente complejo no es ya solo las células cancerosas sino todo el microambiante en que están inmersas.
Exacto, Juan. Gracias.